
Что такое гельминты и какие виды паразитических червей существуют. Как устроены и развиваются сосальщики, ленточные и круглые черви. Каковы особенности жизненных циклов основных групп гельминтов. Как происходит заражение человека различными видами паразитических червей.
Общая характеристика гельминтов
Гельминты — это общее название паразитических червей, вызывающих заболевания у человека и животных. К основным группам гельминтов относятся:
- Сосальщики (трематоды)
- Ленточные черви (цестоды)
- Круглые черви (нематоды)
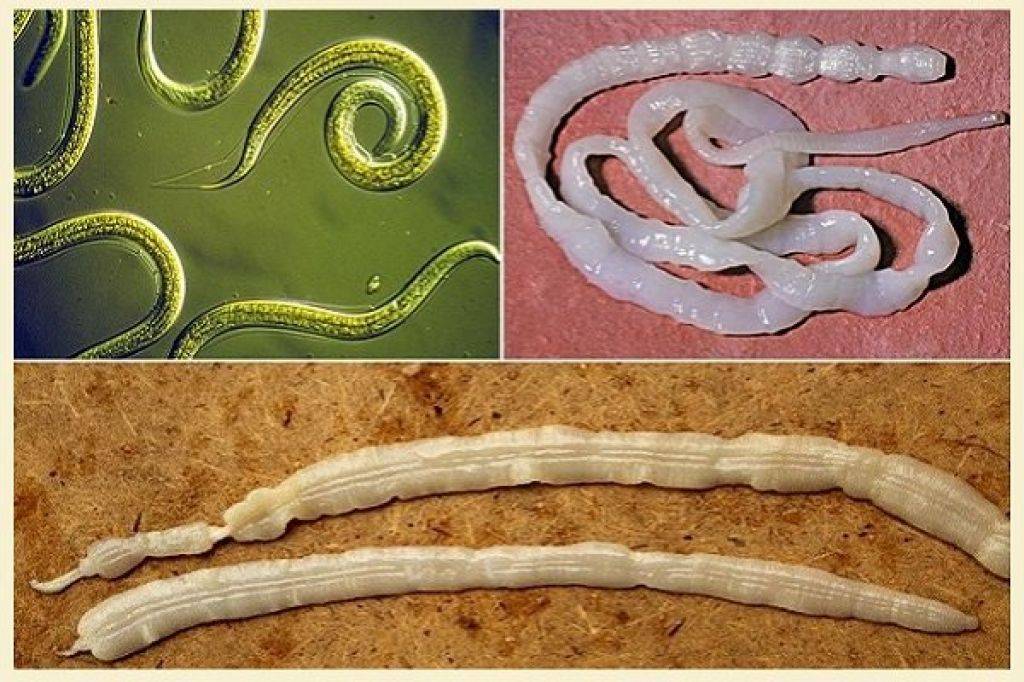
Гельминты характеризуются удлиненным телом и отсутствием конечностей. В процессе эволюции они приспособились к паразитическому образу жизни в организме хозяина. Большинство видов имеют сложные жизненные циклы со сменой нескольких хозяев.
Строение и особенности сосальщиков (трематод)
Сосальщики имеют плоское листовидное тело длиной от нескольких миллиметров до 7-8 см. Основные особенности их строения:
- Наличие ротовой и брюшной присосок для фиксации в организме хозяина
- Отсутствие полости тела, органы погружены в паренхиму
- Наличие кожно-мускульного мешка
- Развитый пищеварительный тракт с разветвленным кишечником
- Выделительная система протонефридиального типа
- Большинство видов — гермафродиты
Яйца сосальщиков имеют крышечку и выделяются с фекалиями, мочой или мокротой хозяина. Жизненный цикл включает несколько стадий развития:
- Яйцо
- Мирацидий — личинка, заражающая промежуточного хозяина (моллюска)
- Спороциста или редия — стадии бесполого размножения в моллюске
- Церкария — хвостатая личинка, покидающая моллюска
- Метацеркария — инцистированная стадия, заразная для окончательного хозяина
- Взрослая особь
Строение и особенности ленточных червей (цестод)
Ленточные черви имеют лентовидное сегментированное тело длиной от нескольких миллиметров до 10-12 метров. Характерные черты цестод:
- Тело состоит из головки (сколекса), шейки и цепочки члеников (стробилы)
- На сколексе расположены органы фиксации — присоски, крючья
- Отсутствие пищеварительной системы, питание через покровы тела
- Каждый членик содержит полный набор половых органов
- Гермафродиты
Жизненный цикл ленточных червей включает следующие стадии:

- Яйцо, содержащее онкосферу (зародыш с крючьями)
- Личинка в промежуточном хозяине (финна, цистицерк, ценур и др.)
- Взрослый червь в окончательном хозяине
Строение и особенности круглых червей (нематод)
Нематоды имеют нитевидное или веретеновидное тело, покрытое плотной кутикулой. Основные черты их организации:
- Наличие первичной полости тела
- Кожно-мускульный мешок из одного слоя продольных мышц
- Пищеварительная система в виде прямой трубки
- Нервная система представлена нервными стволами и кольцом вокруг глотки
- Раздельнополые, выражен половой диморфизм
Жизненный цикл нематод включает следующие стадии:
- Яйцо
- 4 личиночные стадии с линьками
- Взрослая особь
Особенности заражения человека различными гельминтами
Заражение сосальщиками
Основные пути заражения человека трематодами:
- Употребление в пищу сырой, недостаточно термически обработанной рыбы, содержащей метацеркарии (описторхоз, клонорхоз)
- Проникновение церкарий через кожу при контакте с зараженной водой (шистосомозы)
- Поедание водных растений с прикрепленными метацеркариями (фасциолез)
Заражение ленточными червями
Человек может заразиться цестодами следующими путями:
- Употребление недостаточно проваренного или прожаренного мяса, содержащего финны (тениаринхоз, тениоз)
- Случайное проглатывание яиц с загрязненной пищей или водой (эхинококкоз, альвеококкоз)
- Употребление сырой рыбы, содержащей плероцеркоиды (дифиллоботриоз)
Заражение круглыми червями
Основные способы заражения человека нематодами:
- Проглатывание яиц с загрязненной почвой, водой, овощами (аскаридоз, трихоцефалез)
- Активное внедрение личинок через кожу (анкилостомидозы)
- Употребление недостаточно термически обработанного мяса (трихинеллез)
- Укусы кровососущих насекомых — переносчиков микрофилярий (филяриозы)
Патогенное действие гельминтов на организм человека
Паразитические черви оказывают комплексное негативное воздействие на организм хозяина:
- Механическое повреждение тканей и органов
- Нарушение процессов пищеварения и всасывания питательных веществ
- Токсико-аллергическое действие продуктов жизнедеятельности гельминтов
- Снижение иммунитета
- Истощение организма
- Анемия при кровососущих видах
- Закупорка протоков внутренних органов
Диагностика гельминтозов
Для выявления паразитарных заболеваний используют следующие методы:

- Копроовоскопия — обнаружение яиц гельминтов в кале
- Серологические реакции — выявление антител к паразитам
- Иммуноферментный анализ (ИФА)
- Полимеразная цепная реакция (ПЦР)
- Инструментальные методы — УЗИ, КТ, МРТ
- Биопсия тканей
Профилактика гельминтозов
Основные меры профилактики заражения паразитическими червями включают:
- Соблюдение правил личной гигиены
- Тщательное мытье овощей, фруктов, зелени
- Употребление только термически обработанного мяса и рыбы
- Использование репеллентов для защиты от насекомых-переносчиков
- Регулярное обследование на гельминтозы
- Профилактическая дегельминтизация домашних животных
- Санитарно-просветительская работа среди населения
Современные подходы к лечению гельминтозов
Терапия паразитарных заболеваний включает следующие направления:
- Этиотропная терапия — применение антигельминтных препаратов
- Патогенетическая терапия — устранение последствий паразитарной инвазии
- Иммуномодулирующая терапия
- Симптоматическое лечение
- Энтеросорбция для выведения токсинов
- Диетотерапия
Выбор конкретной схемы лечения зависит от вида возбудителя, интенсивности инвазии, наличия осложнений и сопутствующих заболеваний. После окончания терапии обязательно проводится контрольное обследование для подтверждения эффективности лечения.

Заключение
Гельминтозы остаются актуальной проблемой здравоохранения во многих странах мира. Знание особенностей строения, жизненных циклов и путей заражения различными видами паразитических червей необходимо для разработки эффективных мер профилактики и лечения этих заболеваний. Комплексный подход, включающий санитарно-гигиенические мероприятия, своевременную диагностику и рациональную терапию, позволяет успешно бороться с гельминтными инвазиями.
Как защитить домашних питомцев от гельминтов / Новости города / Сайт Москвы
Здравоохранение
Комитет ветеринарии напоминает о необходимости профилактики гельминтозов у животных.Гельминтозы – группа заболеваний животных и людей, которые вызваны присутствием в организме различных паразитов. Гельминтозами болеют все виды домашних и сельскохозяйственных животных. Некоторые виды гельминтов могут передаваться людям от домашних питомцев. Особую опасность для животных и человека представляют такие гельминтозы, как эхинококкоз, альвеококкоз, описторхоз, аскаридоз, токсокароз, анкилостомоз, дирофиляриоз и др. Попадая в организм человека, паразиты могут жить и развиваться в нем до нескольких лет, нанося ущерб здоровью.
Домашние животные могут заразиться гельминтами при контакте с другими животными, через воду, мясо или рыбу и даже при укусах насекомых. Щенки и котята могут получить заболевание от матери при рождении и кормлении.
Симптомы глистной инвазии у домашних питомцев очень разнообразны: изменение аппетита, истощение, снижение иммунитета, аллергические реакции, нарушения в работе желудочно-кишечного тракта и другие. Возможно бессимптомное течение болезни.
Чтобы определить наличие глистной инвазии в организме животного необходимо обратиться в ветеринарную клинику для сдачи анализов. Основные материалы для лабораторной диагностики гельминтозов — кал и кровь, но иногда могут потребоваться и дополнительные обследования
Лечение гельминтозов у животных проводится специальными антигельминтными препаратами под контролем ветеринарного врача. При сильной степени инвазии дегельминтизацию проводят два раза с интервалом 10-14 дней. Для контроля проведенного лечения требуется повторная сдача анализов.
С целью профилактики из рациона животных стоит исключить сырые продукты, не позволять им пить воду из луж и открытых водоемов. Животным, которых содержат в помещении без выгула на улице, рекомендуется давать антигельминтные препараты не реже двух раз в год, а животным, регулярно бывающим на улице, рекомендуется проводить профилактическую дегельминтизацию каждые четыре месяца.
Чтобы избежать заражения гельминтами от домашних животных владельцам животных нужно обязательно соблюдать правила личной гигиены и меры профилактики.
Получить консультацию о методах профилактики гельминтозов у животных и оказании ветеринарной помощи вы можете в государственных ветеринарных учреждениях города Москвы. Контактная информация размещена на портале открытых данных Правительства Москвы в разделе «Центры бесплатной вакцинации против бешенства».
С целью удобства и экономии своего времени вы можете предварительно записаться на прием к ветеринарному специалисту через портал mos.ru.
Теги
Комитет ветеринарии города Москвы государственные ветеринарные клиники профилактика гельминтозов у животных
Сферы
Здравоохранение
Департаменты
Комитет ветеринарии города Москвы
Все новости
Дронтал для кошек от гельминтов (2таб) элипсоидной формы (42676)
ОПИСАНИЕДронтал (Drontal) — таблетки для орального применения для дегельминтизации взрослых кошек. По внешнему виду препарат представляет собой вязкую жидкость светло-желтого цвета. Таблетка белого цвета, на разломе желтоватая, с разделительной бороздкой по центру, массой 350 мг.
По внешнему виду препарат представляет собой вязкую жидкость светло-желтого цвета. Таблетка белого цвета, на разломе желтоватая, с разделительной бороздкой по центру, массой 350 мг.
В своем составе содержит в качестве действующих веществ 230 мг пирантела эмбоната и 20 мг празиквантела, а также вспомогательные компоненты.
ФАРМАКОЛОГИЧЕСКИЕ СВОЙСТВААктивно действующие компоненты дронтала празиквантел и пирантела эмбонат проявляют синергизм действия и губительно действуют против круглых и ленточных гельминтов, в т. ч. Toxocara cati (mystax), Ancylostoma tubaeforme, Ancylostoma braziliense, Uncinaria stenocephala, Echinococcus multilocularis, Dipylidium caninum, Taenia spp., Mesocestoides spp., паразитирующих у кошек. Механизм действия празиквантела заключается в деполяризации нейромышечных ганглиоблокаторов, нарушении транспорта глюкозы и микротубулярной функции у цестод, что приводит к нарушению мышечной иннервации, параличу и гибели паразита.
Празиквантел быстро всасывается в кишечнике и распределяется во внутренних органах и тканях, выводится из организма с мочой в основном в метаболизированной форме. Пирантела эмбонат оказывает выраженное нематодоцидное действие, вызывая повышение проницаемости клеточных мембран, необратимый паралич и контрактуру мышц у нематод, что приводит к гибели и выделению паразитов из желудочно-кишечного тракта. Пирантела эмбонат частично всасывается в кишечнике, быстро метаболизируется и выводится из организма в основном с фекалиями. Дронтал по степени воздействия на организм относится к малоопасным веществам (4 класс опасности по ГОСТ 12.1.007-76), в рекомендуемых дозах не оказывает эмбриотоксического, тератогенного и сенсибилизирующего действия. Хорошо переносится кошками разных пород и возраста.
ПОКАЗАНИЯНазначают кошкам с профилактической и лечебной целью при нематодозах (токсокароз, анкилостомоз, унцинариоз) и цестодозах (тениоз, дипилидиоз, эхинококкоз, мезоцестоидоз).
Дронтал вводят кошкам внутрь индивидуально, однократно в утреннее кормление с небольшим количеством корма (в куске колбасы, мяса, с фаршем). В случае отказа животного от корма с препаратом, а также котятам дронтал вводят принудительно на корень языка или в виде водной суспензии с помощью шприца и катетера. Препарат задают в дозе из расчета 1 таблетка на 4 кг массы животного, что соответствует 5 мг празиквантела и 57,5 мг пирантела эмбоната на 1 кг веса животного. Предварительной голодной диеты и применения слабительных средств перед дегельминтизацией не требуется. С профилактической целью дегельминтизацию кошек проводят ежеквартально, а также перед вакцинацией и случкой.
ПОБОЧНЫЕ ДЕЙСТВИЯПобочных явлений и осложнений при применении дронтала в соответствии с настоящей инструкцией, как правило, не наблюдается. При повышенной индивидуальной чувствительности животного к активным компонентам препарата возможно появление повышенной саливации, диареи или рвоты, указанные симптомы кратковременны, самопроизвольно исчезают и не требуют применения лекарственных средств.
Повышенная индивидуальная чувствительность к компонентам препарата. Не следует применять котятам моложе 3-недельного возраста и беременным кошкам в первые две трети периода беременности.
ОСОБЫЕ УКАЗАНИЯОсобые меры предосторожности не предусмотрены.
СРОК ГОДНОСТИ И ХРАНЕНИЯС предосторожностью по списку Б. В защищенном от света и влаги, недоступном для детей и животных месте при температуре от 0 до 25 °С. Срок годности — 5 лет.
УПАКОВКАВыпускают Дронтал расфасованным по 2 таблетки в блистерах из алюминиевой фольги, упакованных в картонные коробки.
Гельминты: строение, классификация, рост и развитие – медицинская микробиология
Общие понятия
Гельминты – червеобразные паразиты. Клинически значимые группы разделены в соответствии с их общей внешней формой и органом-хозяином, в котором они обитают. Существуют как гермафродиты, так и обоеполые виды. Окончательная классификация основана на внешней и внутренней морфологии яиц, личинок и взрослых стадий.
Окончательная классификация основана на внешней и внутренней морфологии яиц, личинок и взрослых стадий.
Сосальщики (трематоды)
Взрослые двуустки представляют собой листообразных плоских червей. Выступающие ротовые и брюшные присоски помогают удерживать положение на месте. Сосальщики гермафродиты, за исключением двуполых кровяных трематод. Жизненный цикл включает промежуточного хозяина улитки.
Ленточные черви (цестоды)
Взрослые ленточные черви представляют собой удлиненные, сегментированные, гермафродитные плоские черви, обитающие в просвете кишечника. Личиночные формы, кистозные или солидные, населяют внекишечные ткани.
Круглые черви (нематоды)
Взрослые и личиночные круглые черви — двуполые, цилиндрические черви. Они обитают в кишечном и внекишечном отделах.
Введение
Гельминты — это общий термин, обозначающий червей. Гельминты — беспозвоночные, характеризующиеся удлиненным, плоским или круглым телом. В медицинских схемах плоские черви или платигельминты (platy от греческого корня, означающего «плоский») включают сосальщиков и ленточных червей. Круглые черви — это нематоды (немато от греческого корня, означающего «нить»). Эти группы подразделяются для удобства в соответствии с органом хозяина, в котором они обитают, например, легочные двуустки, внекишечные ленточные черви и кишечные аскариды. В этой главе рассматриваются строение и развитие трех основных групп гельминтов.
Круглые черви — это нематоды (немато от греческого корня, означающего «нить»). Эти группы подразделяются для удобства в соответствии с органом хозяина, в котором они обитают, например, легочные двуустки, внекишечные ленточные черви и кишечные аскариды. В этой главе рассматриваются строение и развитие трех основных групп гельминтов.
Гельминты развиваются в стадии яйца, личинки (молодости) и взрослой особи. дает названия, применяемые к различным личиночным гельминтам. Знание различных стадий их роста и развития является основой для понимания эпидемиологии и патогенеза гельминтозов, а также для диагностики и лечения больных, являющихся носителями этих паразитов. Вклады различных стадий в заболевание перечислены в .
Таблица 86-1
Распространенные личиночные формы гельминтов, встречающиеся у людей.
Таблица 86-2
Стадии гельминтов, обычно ответственных за патологические изменения в организме человека.
Платигельминты и нематоды, поражающие человека, имеют схожие анатомические особенности, которые отражают общие физиологические потребности и функции. Наружный покров гельминтов — кутикула или тегумент. Выдающимися внешними структурами трематод и цестод являются вертлужные впадины (присоски) или ботрии (ложные присоски). Самцы нематод некоторых видов обладают дополнительными половыми органами, являющимися внешними модификациями кутикулы. Опытный наблюдатель может идентифицировать пищеварительную, выделительную и репродуктивную системы изнутри. Ленточные черви уникальны отсутствием пищеварительного канала. Этот недостаток означает, что питательные вещества должны всасываться через покровы. Кровяные сосальщики и нематоды обоеполые. Все другие виды трематод и ленточных червей, заражающих человека, являются гермафродитами.
Наружный покров гельминтов — кутикула или тегумент. Выдающимися внешними структурами трематод и цестод являются вертлужные впадины (присоски) или ботрии (ложные присоски). Самцы нематод некоторых видов обладают дополнительными половыми органами, являющимися внешними модификациями кутикулы. Опытный наблюдатель может идентифицировать пищеварительную, выделительную и репродуктивную системы изнутри. Ленточные черви уникальны отсутствием пищеварительного канала. Этот недостаток означает, что питательные вещества должны всасываться через покровы. Кровяные сосальщики и нематоды обоеполые. Все другие виды трематод и ленточных червей, заражающих человека, являются гермафродитами.
За немногими исключениями, взрослые двуустки, цестоды и нематоды производят яйца, которые выделяются с выделениями или выделениями хозяина. Различные стадии и их уникальные характеристики будут рассмотрены более подробно по мере рассмотрения каждой основной группы гельминтов.
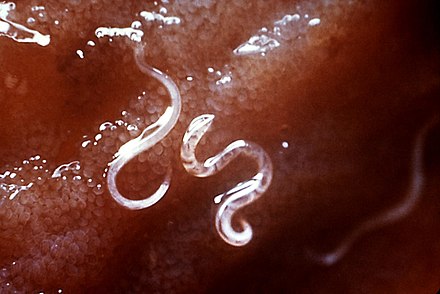
Сосальщики (трематоды)
Структура трематод представлена в и . Уплощенное в дорсовентральном направлении тело, билатеральная симметрия и четко выраженный передний конец — признаки платигельминтов вообще и трематод в частности. Сосальщики листовидные, длиной от нескольких миллиметров до 7-8 см. Тегумент морфологически и физиологически сложен. Сосальщики имеют ротовую присоску вокруг рта и брюшную присоску или вертлужную впадину, которую можно использовать для прикрепления к тканям хозяина. Полость тела отсутствует. Органы погружены в специализированную соединительную ткань или паренхиму. Слои соматических мышц пронизывают паренхиму и прикрепляются к тегументу.
Уплощенное в дорсовентральном направлении тело, билатеральная симметрия и четко выраженный передний конец — признаки платигельминтов вообще и трематод в частности. Сосальщики листовидные, длиной от нескольких миллиметров до 7-8 см. Тегумент морфологически и физиологически сложен. Сосальщики имеют ротовую присоску вокруг рта и брюшную присоску или вертлужную впадину, которую можно использовать для прикрепления к тканям хозяина. Полость тела отсутствует. Органы погружены в специализированную соединительную ткань или паренхиму. Слои соматических мышц пронизывают паренхиму и прикрепляются к тегументу.
Рисунок 86-1
Структура лап. (А) Гермафродитная двуустка. (B) Бисексуальная двуустка. (С изменениями из Hunter GW, Swartzwelder JC, Clyde DF: A Manual of Tropical Medicine. 5th Ed. WB Saunders, Philadelphia, 1976, с разрешения.)
Во всех циклах в качестве промежуточных хозяев участвуют улитки. Гермафродитные двуустки —
Clonorchis sinensis, Fasiolopsis buski, Paragonimys westermani, и Heterophytes heterpphyes . Метацеркарии заразны для человека. Двуполые двуустки: (далее…)
Метацеркарии заразны для человека. Двуполые двуустки: (далее…)Сосальщики имеют хорошо развитый пищеварительный тракт с мускулистыми глоткой и пищеводом. Кишечник обычно представляет собой разветвленную трубку (могут присутствовать вторичные и третичные ветви), состоящую из одного слоя эпителиальных клеток. Основные ветви могут заканчиваться слепо или открываться в экскреторный пузырек. Выделительный пузырек также принимает два основных боковых собирательных протока выделительной системы протонефридиального типа с пламенными клетками. Пламенная клетка представляет собой полую конечную экскреторную клетку, содержащую бьющуюся (похожую на пламя) группу ресничек. Эти клетки, закрепленные в паренхиме, направляют тканевый фильтрат через каналы в два главных собирательных протока.
За исключением кровяных сосальщиков, трематоды являются гермафродитами, имеющими как мужские, так и женские репродуктивные органы у одного и того же человека. Мужской орган обычно состоит из двух семенников с добавочными железами и протоками, ведущими к циррусу или эквиваленту полового члена, который простирается в общий половой атриум. Женская гонада состоит из одного яичника с семеприемником и желточниками, или желточными железами, которые соединяются с яйцеводом по мере его расширения в оотип. Трубчатая матка отходит от оотипа и открывается в половой атриум. Происходит как само-, так и перекрестное оплодотворение. Компоненты яйца собираются в оотипе. Яйца проходят через матку в генитальный атриум и выходят вентрально через генитальное отверстие. Яйца двуустки, кроме шистосом, с крышечкой (крышечкой).
Женская гонада состоит из одного яичника с семеприемником и желточниками, или желточными железами, которые соединяются с яйцеводом по мере его расширения в оотип. Трубчатая матка отходит от оотипа и открывается в половой атриум. Происходит как само-, так и перекрестное оплодотворение. Компоненты яйца собираются в оотипе. Яйца проходят через матку в генитальный атриум и выходят вентрально через генитальное отверстие. Яйца двуустки, кроме шистосом, с крышечкой (крышечкой).
Кровяные двуустки или шистосомы — единственные двуполые двуустки, заражающие людей (). Хотя полы раздельны, общее строение тела такое же, как у трематод-гермафродитов. Внутри окончательного хозяина самцы и самки червей обитают в просвете кровеносных сосудов и находятся в тесной физической связи. Самка лежит внутри покровной складки, гинекофорного канала, на вентральной поверхности самца. Медицински важные двуустки относятся к таксономической категории Digenea. Цикл развития этой группы сосальщиков требует как минимум двух хозяев, один из которых является промежуточным хозяином улитки. В зависимости от вида могут быть задействованы другие промежуточные хозяева для сохранения личиночной формы, заражающей окончательного хозяина-человека.
В зависимости от вида могут быть задействованы другие промежуточные хозяева для сохранения личиночной формы, заражающей окончательного хозяина-человека.
Сосальщики проходят несколько личиночных стадий, каждая из которых имеет определенное имя, прежде чем достичь зрелости. Принимая во внимание различия между видами (см.), обобщенный жизненный цикл дигенетических трематод протекает следующим образом. Яйца попадают в фекалии, мочу или мокроту человека и попадают в водную среду. Из яиц вылупляются реснитчатые личинки или мирацидии, которые либо проникают внутрь, либо поедаются промежуточным хозяином улиткой. В редких случаях наземные улитки могут служить промежуточными хозяевами. Мешковидная стадия спороцисты или редии развивается из мирацидия в тканях улитки.
Спороциста дает начало либо редии, либо стадии дочерней спороцисты. В свою очередь, из редии или дочерней спороцисты церкарии развиваются бесполым путем и мигрируют из тканей улитки во внешнюю среду, обычно водную.
Церкарии, которые могут иметь плавательный хвост, в дальнейшем развиваются одним из трех способов. Они либо проникают в окончательного хозяина и превращаются непосредственно во взрослых особей, либо проникают во второго промежуточного хозяина и развиваются как инцистированные метацеркарии, либо инцистируют на субстрате, например в растительности, и развиваются там как метацеркарии. Когда метацеркарная киста проглатывается, переваривание кисты высвобождает незрелую двуустка, которая мигрирует в определенное место органа и развивается во взрослую червь.
Они либо проникают в окончательного хозяина и превращаются непосредственно во взрослых особей, либо проникают во второго промежуточного хозяина и развиваются как инцистированные метацеркарии, либо инцистируют на субстрате, например в растительности, и развиваются там как метацеркарии. Когда метацеркарная киста проглатывается, переваривание кисты высвобождает незрелую двуустка, которая мигрирует в определенное место органа и развивается во взрослую червь.
Ленточные черви (цестоды)
Являясь представителями платигельминтов, цестоды, или ленточные черви, обладают многими основными структурными характеристиками сосальщиков, но также обнаруживают поразительные отличия. показаны общие черты строения и развития ленточных червей.
Рисунок 86-3
Строение ленточных червей. (С изменениями из Jeffery HC, Leach RM: Atlas of Medical Helminthology and Protozoology. Churchill Livingstone, Edinburgh, 1968, с разрешения.) называются проглоттидами. Ленточные черви имеют длину от 2–3 мм до 10 м и могут иметь от трех до нескольких тысяч сегментов.
Анатомически цестоды делятся на сколекс, или головку, на которой расположены органы прикрепления, шейку, являющуюся областью пролиферации сегментов, и цепочку проглоттид, называемую стробилой. Стробила удлиняется по мере того, как в области шейки образуются новые проглоттиды. Ближайшие к шее сегменты незрелые (половые органы не полностью развиты), а более задние — зрелые. Конечные сегменты беременны, наиболее заметной особенностью является наполненная яйцами матка.
Сколекс содержит головной ганглий, или «мозг», нервной системы ленточного червя. Внешне сколекс характеризуется удерживающими органами. В зависимости от вида эти органы состоят из хоботка, ботрии или вертлужной впадины. Ростеллум представляет собой выдвижную конусообразную структуру, расположенную на переднем конце сколекса и у некоторых видов вооруженную крючками. Ботрии представляют собой длинные, узкие, слабомускулистые бороздки, характерные для псевдофиллидовых цепней. Вертлужные впадины (присоски, как у дигенетических трематод) характерны для циклофиллидных ленточных червей.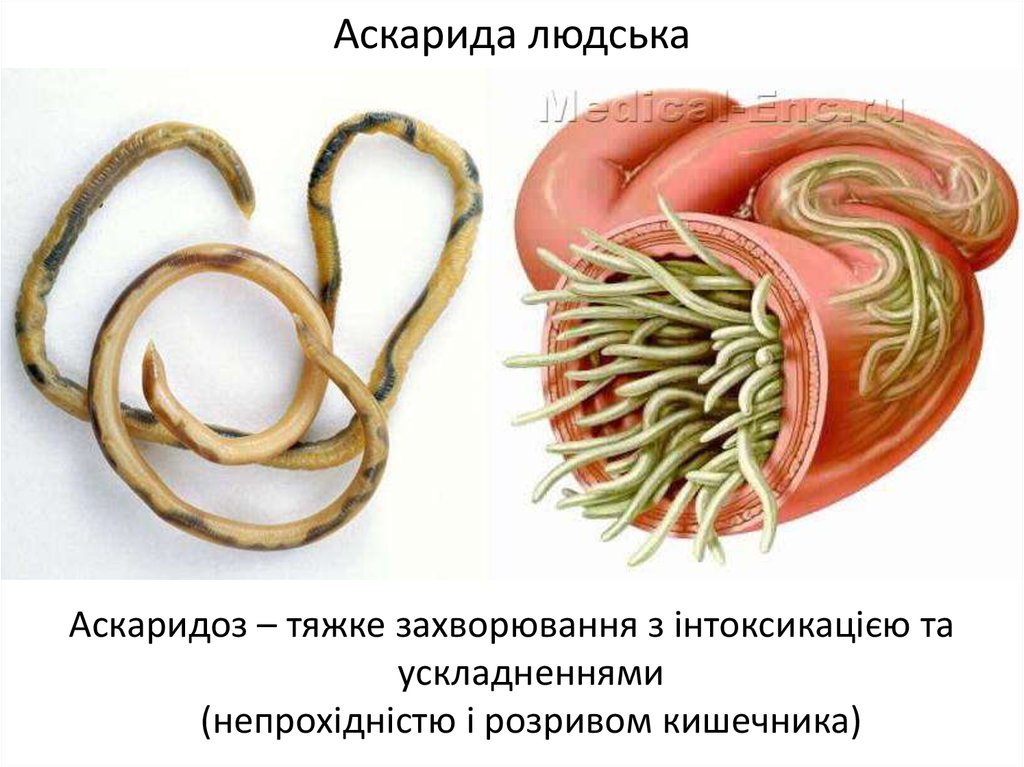 Дифференциальные признаки псевдофиллидных и циклофиллидных ленточных червей перечислены в . Большинство ленточных червей человека относятся к циклофиллидам.
Дифференциальные признаки псевдофиллидных и циклофиллидных ленточных червей перечислены в . Большинство ленточных червей человека относятся к циклофиллидам.
Таблица 86-3
Различия между псевдофиллидными и циклофиллидными цепнями.
Характерной чертой взрослых ленточных червей является отсутствие пищеварительного канала, что интересно, поскольку все эти взрослые черви обитают в тонком кишечнике. Отсутствие пищеварительного тракта означает, что вещества попадают к лентецу через покровы. Эта структура хорошо приспособлена к транспортным функциям, так как покрыта многочисленными микроворсинками, напоминающими те, что выстилают просвет кишечника млекопитающих. Выделительная система относится к типу пламенных клеток.
Цестоды являются гермафродитами, каждая проглоттида имеет мужские и женские репродуктивные системы, подобные таковым у дигенетических сосальщиков. Однако ленточные черви отличаются от сосальщиков механизмом откладывания яиц. Яйца псевдофиллидовых цепней выходят через пору матки в центре вентральной поверхности, а не через генитальный атриум, как у сосальщиков. У циклофиллидных цепней женская система включает матку без маточной поры (). Таким образом, циклофиллидные яйца высвобождаются только тогда, когда ленточные черви выделяют беременные проглоттиды в кишечник. Некоторые проглоттиды распадаются, высвобождая яйца, которые выделяются с фекалиями, тогда как другие проглоттиды выходят целыми.
У циклофиллидных цепней женская система включает матку без маточной поры (). Таким образом, циклофиллидные яйца высвобождаются только тогда, когда ленточные черви выделяют беременные проглоттиды в кишечник. Некоторые проглоттиды распадаются, высвобождая яйца, которые выделяются с фекалиями, тогда как другие проглоттиды выходят целыми.
Яйца псевдофиллидных ленточных червей с крышечкой, а циклофиллидных — нет. Однако яйца всех ленточных червей на той или иной стадии развития содержат зародыш или онкосферу. Онкосфера псевдофиллидовых цепней снаружи реснитчатая и называется корацидием. Корацидий развивается в процеркоидную стадию у своего первого непосредственного хозяина из микроракообразных, а затем в личинку плероцеркоида у своего следующего промежуточного хозяина, которым является позвоночное. Личинка плероцеркоида развивается во взрослую особь у окончательного (окончательного) хозяина. Онкосфера циклофиллидных цепней в зависимости от вида развивается в личинку цистицерка, личинку цистицеркоида, личинку ценуруса или эхинококковую личинку (цисту) у конкретных промежуточных хозяев. Эти личинки, в свою очередь, превращаются во взрослых особей в окончательном хозяине. иллюстрирует эти личиночные формы и репрезентативные жизненные циклы.
Эти личинки, в свою очередь, превращаются во взрослых особей в окончательном хозяине. иллюстрирует эти личиночные формы и репрезентативные жизненные циклы.
Рисунок 86-4
Обобщенный жизненный цикл ленточных червей. Hymenolepsis nana, Hdiminuta, Taenia saginata, T solium, Diphyllobothrium latum, Dipylidium craninum . Обратите внимание на зародыши гексаканта. личинка цистицерка у коровы и свиньи; личинка процеркоида у копепод, плероцеркоид (sparganum) (подробнее…)
Круглые черви (нематоды)
показано строение нематод. В отличие от платигельминтов, нематоды имеют цилиндрическую, а не уплощенную форму; отсюда и общее название аскариды. Стенка тела состоит из наружной кутикулы, имеющей неклеточную, химически сложную структуру, тонкой гиподермы и мускулатуры. Кутикула у некоторых видов имеет продольные гребни, называемые крыльями. Бурса, лоскутное расширение кутикулы на заднем конце некоторых видов самцов нематод, используется для захвата самки во время копуляции.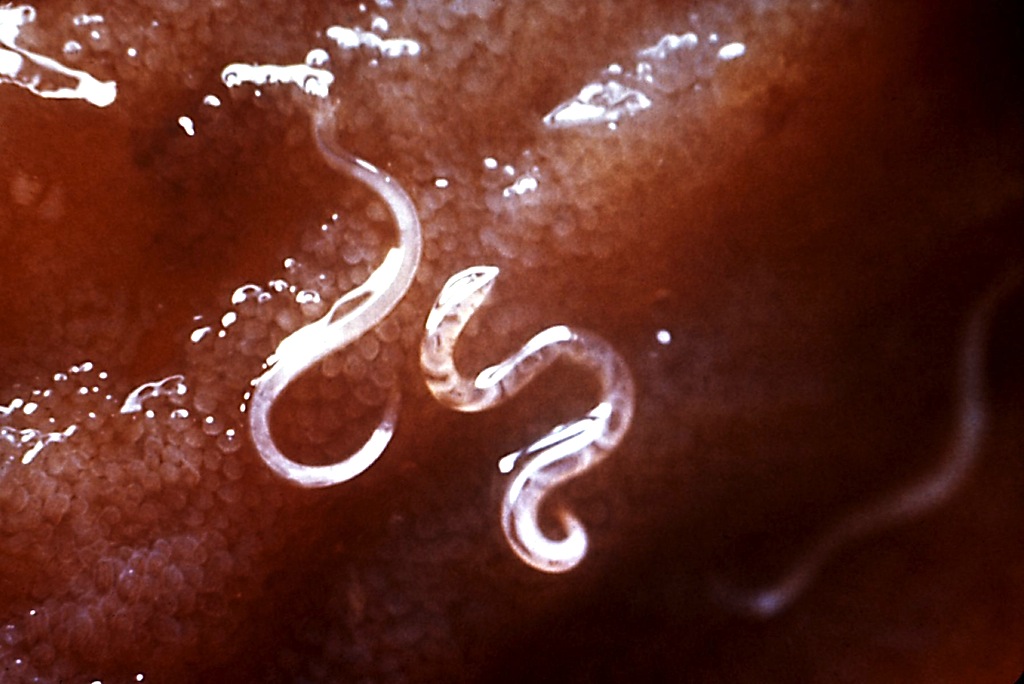
Рисунок 86-5
Структура нематод. (Женщина. (Б) Мужчина. Поперечные срезы через среднюю область самки червя (C) и через область пищевода (D). (С изменениями из Lee DL: The Physiology of Nematodes. Oliver and Boyd, Edinburgh, 1965, с разрешения.) (подробнее…)
Клеточная гиподерма выпячивается в полость тела или псевдоцелом, образуя четыре продольных тяжа — дорсальный, брюшной и два боковых шнура, которые можно увидеть на поверхности как боковые линии. Ядра гиподермы располагаются в области тяжей. Соматическая мускулатура, лежащая под гиподермой, представляет собой один слой гладкомышечных клеток. При просмотре в поперечном сечении можно увидеть, что этот слой разделен гиподермальными тяжами на четыре зоны. Мускулатура иннервируется отростками мышечных клеток к нервным стволам, идущим кпереди и кзади от ганглиозных клеток, окружающих среднюю часть пищевода.
Пространство между мышечным слоем и внутренностями представляет собой псевдоцелом, лишенный мезотелиальной выстилки. Эта полость содержит жидкость и от двух до шести фиксированных клеток (целомоцитов), которые обычно связаны с продольными тяжами. Функция этих клеток неизвестна.
Эта полость содержит жидкость и от двух до шести фиксированных клеток (целомоцитов), которые обычно связаны с продольными тяжами. Функция этих клеток неизвестна.
Пищеварительный тракт круглых червей полный, с ротовым и анальным отверстиями. Рот окружен губами с чувствительными сосочками (щетинками). Пищевод, заметная черта нематод, представляет собой мышечную структуру, которая перекачивает пищу в кишечник; она отличается по форме у разных видов.
Кишечник представляет собой трубчатую структуру, состоящую из одного слоя столбчатых клеток с заметными микроворсинками на поверхности просвета.
Выделительная система некоторых нематод состоит из выделительной железы и поры, расположенной вентрально в средней части пищевода. У других нематод эта структура втягивается в расширения, дающие начало более сложной трубчатой выделительной системе, обычно Н-образной формы, с двумя передними и двумя задними конечностями, расположенными в боковых тяжах. Считается, что железистые клетки и трубки служат абсорбирующими органами, собирая отходы из псевдоцелома и участвующие в осморегуляции.
Нематоды обычно обоеполые. Самцы обычно меньше самок, имеют изогнутый задний конец и обладают (у некоторых видов) копулятивными структурами, такими как спикулы (обычно две), бурса или и то, и другое. У самцов есть один или (в некоторых случаях) два семенника, которые лежат на свободном конце извитой или загнутой трубки, ведущей в семенной пузырь и, в конечном итоге, в клоаку.
Женская система также трубчатая и обычно состоит из загнутых яичников. Каждый яичник сплошной, с яйцеводом и трубчатой маткой. Матки соединяются, образуя влагалище, которое, в свою очередь, открывается наружу через вульву.
Совокупление самки и самца нематоды необходимо для оплодотворения, за исключением представителей рода Strongyloides, , у которых происходит партеногенетическое развитие (т. е. развитие неоплодотворенной яйцеклетки в новую особь). Некоторые данные указывают на то, что половые аттрактанты (феромоны) играют роль в гетеросексуальном спаривании. Во время совокупления сперма попадает в вульву самки. Сперматозоид попадает в яйцеклетку, и зигота выделяет мембрану для оплодотворения. Эта оболочка постепенно утолщается, образуя хитиновую оболочку. Вторая мембрана под скорлупой делает яйцо непроницаемым практически для всех веществ, кроме углекислого газа и кислорода. У некоторых видов выделяется третья белковая оболочка, когда яйцеклетка проходит по матке вдоль стенки матки и откладывается вне скорлупы. Большинство нематод, паразитирующих у человека, откладывают яйца, которые при мочеиспускании содержат либо нерасщепленную зиготу, группу бластомеров, либо полностью сформировавшуюся личинку. Некоторые нематоды, такие как филярии и Trichinella Spiralis производят личинок, которые откладываются в тканях хозяина.
Сперматозоид попадает в яйцеклетку, и зигота выделяет мембрану для оплодотворения. Эта оболочка постепенно утолщается, образуя хитиновую оболочку. Вторая мембрана под скорлупой делает яйцо непроницаемым практически для всех веществ, кроме углекислого газа и кислорода. У некоторых видов выделяется третья белковая оболочка, когда яйцеклетка проходит по матке вдоль стенки матки и откладывается вне скорлупы. Большинство нематод, паразитирующих у человека, откладывают яйца, которые при мочеиспускании содержат либо нерасщепленную зиготу, группу бластомеров, либо полностью сформировавшуюся личинку. Некоторые нематоды, такие как филярии и Trichinella Spiralis производят личинок, которые откладываются в тканях хозяина.
Процесс развития нематод включает стадии яйца, личинки и взрослой особи. За каждой из четырех личиночных стадий следует линька, при которой сбрасывается кутикула. Личинок называют личинками второго возраста после первой линьки и т. д. (). Нематода, образующаяся на пятой стадии, является взрослой. суммирует жизненные циклы нескольких кишечных нематод.
суммирует жизненные циклы нескольких кишечных нематод.
Рисунок 86-6
Стадии развития нематод. (Адаптировано из Lee DL: The Physiology of Nemotodes. Oliver and Boyd, Edinburgh, 19.65, с разрешения.)
Рисунок 86-7
Обобщенный жизненный цикл кишечных нематод.
Ссылки
Ash L, Orihel TC: Паразиты: Руководство по лабораторным процедурам и идентификации. Американское общество клинических патологов, Чикаго, 1987 г. .
Богитш Б.Дж. и Ченг Т.С.: Паразитология человека. У. Б. Сондерс, Филадельфия, 1990 г. .
Castro GA: Трематоды: шистосомоз. p 1710. В Kelly WN (ed): Учебник по внутренним болезням. Дж. Б. Липпинкотт, Филадельфия, 19 лет.89 .
Хантер Г.В., Шварцвелдер Дж.К., Клайд Д.Ф.: Руководство по тропической медицине. 5-е изд. У. Б. Сондерс, Филадельфия, 1976 г. .
Джеффри Х.К., Лич Р.М.: Атлас медицинской гельминтологии и протозоологии. Черчилль Ливингстон, Эдинбург, 1968 год.
 .
.Lee DL: Физиология нематод. Оливер и Бойд, Эдинбург, 1965 год. .
Смит Д.Д.: Физиология трематод. Оливер и Бойд, Эдинбург, 1966 год. .
Шмидт Г.Д., Робертс Л.С.: Основы паразитологии. 3-е изд. Издательство Times Mirror / Mosby College, Сент-Луис, 1985 г. .
Замен V: Атлас медицинской паразитологии. Леа и Фебигер, Филадельфия, 1979 г. .
Уроки гельминтов: чему нас научили гельминты в отношении иммунологии слизистых оболочек
Уроки гельминтов: чему нас научили гельминты в отношении иммунологии слизистых оболочек
Скачать PDF
Скачать PDF
- Редакция
- Джорджия Перона-Райт 1 и
- Генри Дж.
 МакСорли
ORCID: orcid.org/0000-0003-2761-4275 2
МакСорли
ORCID: orcid.org/0000-0003-2761-4275 2
Иммунология слизистых оболочек том 15 , страницы 1049–1051 (2022)Цитировать эту статью
1813 доступов
28 Альтметрический
Детали показателей
Паразитические гельминты являются уникальными патогенами, являясь самыми крупными, наиболее хроническими и, тем не менее, часто наименее патогенными инфекционными агентами, с которыми мы сталкиваемся. Они перемещаются и сохраняются во многих участках слизистой оболочки, и их влияние, экспериментально и клинически, поддерживает многие из наших современных моделей иммунитета слизистой оболочки. Здесь мы представляем специальный сборник недавних статей по иммунологии слизистых оболочек, в которых освещаются ключевые достижения, ставшие возможными благодаря изучению гельминтозов, от открытия новых иммуноэпителиальных клеток до межклеточных взаимодействий, управляющих кишечным иммунитетом, и терапевтических перспектив подавление иммунитета гельминтами. Вместе эти статьи прославляют способность червя дать новое представление о механизмах иммунитета слизистых оболочек, и мы надеемся, что вам понравится читать сборник.
Они перемещаются и сохраняются во многих участках слизистой оболочки, и их влияние, экспериментально и клинически, поддерживает многие из наших современных моделей иммунитета слизистой оболочки. Здесь мы представляем специальный сборник недавних статей по иммунологии слизистых оболочек, в которых освещаются ключевые достижения, ставшие возможными благодаря изучению гельминтозов, от открытия новых иммуноэпителиальных клеток до межклеточных взаимодействий, управляющих кишечным иммунитетом, и терапевтических перспектив подавление иммунитета гельминтами. Вместе эти статьи прославляют способность червя дать новое представление о механизмах иммунитета слизистых оболочек, и мы надеемся, что вам понравится читать сборник.
Уроки гельминтов стали известны очень давно. Паразитические гельминты видны невооруженным глазом и были первыми патогенами, описанными в самом раннем известном медицинском тексте, папирусе Эберса (~ 1500 г. до н.э.), описывающем инфекции человеческого цепня, аскариды, шистосомы, анкилостомы и дракункулеза 1 . Гельминты уже давно связаны с иммунными реакциями типа 2, многие аспекты которых были обнаружены с помощью экспериментальных инфекций (рис. 1). Первыми идентифицированными были эозинофилы и тучные клетки, обнаруженные в конце 1800-х Паулем Эрлихом 9.0190 2 и позднее было показано, что они связаны с паразитарными гельминтозами 3,4 (рис. 1-1). Аналогичным образом, ранние исследования гельминтозов привели Бриджит Огилви и ее команду к описанию «регининоподобных антител» (позже переименованных в IgE) в 1964 г. 1 (рис. 1-2). Когда Мосманн и Коффман первоначально предложили парадигму Th2/Th3, это было подтверждено идентификацией клеток фенотипа Th3 у инфицированных гельминтами мышей 5 (рис. 1-3), в то время как влияние сигнатурных цитокинов Th3 было продемонстрировано в гельминтозы животных с ранним нокаутом по IL-4 и IL-13 6,7,8 . Первоначально считалось, что IL-4 и IL-13 выполняют дублирующие роли в иммунном ответе, пока исследования гельминтозов не выявили доминирующую роль IL-13 в изгнании власоглавов 9 .
Гельминты уже давно связаны с иммунными реакциями типа 2, многие аспекты которых были обнаружены с помощью экспериментальных инфекций (рис. 1). Первыми идентифицированными были эозинофилы и тучные клетки, обнаруженные в конце 1800-х Паулем Эрлихом 9.0190 2 и позднее было показано, что они связаны с паразитарными гельминтозами 3,4 (рис. 1-1). Аналогичным образом, ранние исследования гельминтозов привели Бриджит Огилви и ее команду к описанию «регининоподобных антител» (позже переименованных в IgE) в 1964 г. 1 (рис. 1-2). Когда Мосманн и Коффман первоначально предложили парадигму Th2/Th3, это было подтверждено идентификацией клеток фенотипа Th3 у инфицированных гельминтами мышей 5 (рис. 1-3), в то время как влияние сигнатурных цитокинов Th3 было продемонстрировано в гельминтозы животных с ранним нокаутом по IL-4 и IL-13 6,7,8 . Первоначально считалось, что IL-4 и IL-13 выполняют дублирующие роли в иммунном ответе, пока исследования гельминтозов не выявили доминирующую роль IL-13 в изгнании власоглавов 9 . Позже было показано, что IL-13 является критическим цитокином на поверхности слизистых оболочек, активируя «эпителиальный эскалатор» (увеличение эпителиального обновления) 10 и являясь ключевой частью реакции «плачу и подметаю» (скоординированное увеличение проницаемости эпителия, образование слизи). и перистальтику кишечника), который очищает кишечные паразиты 11 (рис. 1-6). Первые экспериментальные данные, подтверждающие, что слизистый слой является ключевой частью мукозального иммунитета, были также получены при экспериментальных гельминтозах в начале 1980-х годов 12 . Совсем недавно первые исследования альтернативной активации макрофагов 13 (рис. 1-5) и концепции заживления ран как иммунного результата 14 также были основаны на экспериментах по заражению гельминтами. Такие инфекции также показали, что тканевые макрофаги пролиферируют in situ 15 (рис. 1-8) и что Т-фолликулярные хелперные клетки функционально поляризованы 16,17 .
Позже было показано, что IL-13 является критическим цитокином на поверхности слизистых оболочек, активируя «эпителиальный эскалатор» (увеличение эпителиального обновления) 10 и являясь ключевой частью реакции «плачу и подметаю» (скоординированное увеличение проницаемости эпителия, образование слизи). и перистальтику кишечника), который очищает кишечные паразиты 11 (рис. 1-6). Первые экспериментальные данные, подтверждающие, что слизистый слой является ключевой частью мукозального иммунитета, были также получены при экспериментальных гельминтозах в начале 1980-х годов 12 . Совсем недавно первые исследования альтернативной активации макрофагов 13 (рис. 1-5) и концепции заживления ран как иммунного результата 14 также были основаны на экспериментах по заражению гельминтами. Такие инфекции также показали, что тканевые макрофаги пролиферируют in situ 15 (рис. 1-8) и что Т-фолликулярные хелперные клетки функционально поляризованы 16,17 . Наконец, идентификация и характеристика 2 важнейших инициаторов и регуляторов иммунитета типа 2 — врожденных лимфоидных клеток типа 2 (ILC2) 18 (рис. 1-7) и пучковых клеток 19 (рис. 1-10) — оба зависели от использования гельминтозов.
Наконец, идентификация и характеристика 2 важнейших инициаторов и регуляторов иммунитета типа 2 — врожденных лимфоидных клеток типа 2 (ILC2) 18 (рис. 1-7) и пучковых клеток 19 (рис. 1-10) — оба зависели от использования гельминтозов.
1. Идентификация тучных клеток и эозинофилов (конец 1800-х гг.). 2. IgE («реагининоподобные антитела»), индуцируемые при заражении паразитами (1964). 3. Парадигма Th2/Th3: Th2 при бактериальных и Th3 при паразитарных инфекциях (1989). 4. Открытие регуляторных Т-клеток (1995 г.). 5. Парадигма М1/М2, макрофаги М2 («альтернативно активируемые») при паразитарных инфекциях (2000). 6. «Эпителиальный эскалатор» как элемент борьбы с кишечными гельминтами (2005 г.). 7. Характеристика антипаразитарной роли врожденных лимфоидных клеток 2 типа (ILC2) (2010). 8. Пролиферация макрофагов М2 in situ (2011 г.). 9. Метаболический сдвиг при иммунитете 2 типа с гликолиза на метаболизм липидов (2016). 10. Идентификация пучковых клеток как критического типа эпителиальных клеток в инициации антипаразитарного иммунитета (2016). Создано с BioRender.com.
10. Идентификация пучковых клеток как критического типа эпителиальных клеток в инициации антипаразитарного иммунитета (2016). Создано с BioRender.com.
Изображение в полный размер
Сегодня вклад гельминтов в фундаментальное понимание иммунологии слизистых оболочек продолжает увеличиваться. Две исследовательские статьи в этом сборнике описывают диалог между клетками пучка и ILC2: во-первых, показано, что цитокин MIF необходим для активации клеток пучка и ILC2, что имеет решающее значение для быстрого изгнания кишечных гельминтов 20 , а во-вторых, что член семейства SOCS CISH регулирует эту цепь пучка клеток-ILC2, контролируя пороги взаимодействия эпителиальных и иммунных клеток 21 . В обзорной статье этого выпуска Inclan-Rico et al. обсудить текущее понимание межклеточных взаимодействий в инфицированной слизистой оболочке, подчеркнув функцию нетрадиционных иммунных клеток, таких как эпителиальные клетки, нейроны и фибробласты, и утверждая, что связь между гемопоэтическими, стромальными и нервными клетками необходима не только для инициации слизистого иммунитета, но и для его усиления, регуляции и восстановления 22 .
Тема разрешения и репарации продолжена в исследовании, показывающем, что фосфатаза PTPN2 является критическим сигнальным этапом в альтернативной активации макрофагов, а ее отсутствие коррелирует с чрезмерным поражением легких в модели легочной гельминтной инфекции 23 . Иммунитет типа 2 (включая альтернативную активацию макрофагов) связан с особыми метаболическими потребностями, предпочитающими окислительное фосфорилирование аэробному гликолизу, и эти метаболические изменения могут иметь как диетические, так и иммунологические детерминанты (рис. 1-9). Наша коллекция включает интригующее новое исследование активации ILC в кишечнике, которое демонстрирует, что нейропептид, вырабатываемый при употреблении пищи, VIP, взаимодействует с цитокинами-аларминами для мощной активации ILC2 и ILC3 в тканях кишечника 24 . В новом обзоре этого выпуска Michla et al. фокусируется на передовой биологии ILC2 и особенно на метаболическом давлении, которое регулирует активность ILC и последующие иммунные решения 25 .
Одной из особенностей участков слизистой оболочки является постоянное взаимодействие с внешними раздражителями, такими как пища, пыль и комменсальные микроорганизмы. Инфекции слизистых оболочек происходят в контексте этой фоновой стимуляции, а гельминтозы позволили по-новому взглянуть на происходящие взаимодействия. Недавняя статья в нашей коллекции показывает, что бактериальная микробиота также влияет на изгнание гельминтов, регулируя сократительную способность кишечника 26 . Перекрестная регуляция также происходит через иммунную систему, и в двух новых статьях подчеркивается влияние взаимодействия между противоположными реакциями цитокинов. Оба описывают основной ответ IFNγ, присутствующий во время заражения гельминтами, который ограничивает ответ Th3, критический для изгнания паразита 27,28 , и показано, что IL-10 имеет решающее значение для контроля этого IFNγ 28 .
Иммунная регуляция, обнаруженная при заражении гельминтами, сосредоточена в месте заражения, но эти инфекции также выявили дальнодействующую иммунную модуляцию, что способствует нашему пониманию связи между удаленными участками иммунной системы. Гельминты сыграли ключевую роль в описании оси кишечник-легкие, включая раннюю демонстрацию того, что кишечная инфекция может изменить заболевание легких. Связи между другими дистальными участками в настоящее время устанавливаются, и новая захватывающая статья в нашей коллекции раскрывает глубокие изменения в иммунных популяциях в коже во время чисто кишечной гельминтной инфекции 29 . Как тканевая специфичность иммунных ответов, так и связи между отдаленными местами исследуются в новом обзоре Vacca et al. в этом выпуске 30 , подчеркивая большой охват гельминтозов.
Гельминты сыграли ключевую роль в описании оси кишечник-легкие, включая раннюю демонстрацию того, что кишечная инфекция может изменить заболевание легких. Связи между другими дистальными участками в настоящее время устанавливаются, и новая захватывающая статья в нашей коллекции раскрывает глубокие изменения в иммунных популяциях в коже во время чисто кишечной гельминтной инфекции 29 . Как тканевая специфичность иммунных ответов, так и связи между отдаленными местами исследуются в новом обзоре Vacca et al. в этом выпуске 30 , подчеркивая большой охват гельминтозов.
Механизмы иммунной регуляции, обнаруженные при гельминтозах, также стали мощным двигателем терапевтических стратегий, направленных на подавление и смягчение патологических иммунных реакций в различных участках тканей. Вскоре после открытия регуляторных Т-клеток 31 было обнаружено, что паразиты вызывают регуляторные реакции и подавляют иммунитет типа 2 за счет высвобождения иммуномодулирующих продуктов 32 (рис. 1-4). Локе и др. просмотреть последнюю информацию об иммунном ответе человека на паразитических гельминтов, узнать, как последние технологические разработки, такие как исследования на людях, секвенирование отдельных клеток и системы культивирования органоидов, помогли этим исследованиям. Недавние открытия позволяют использовать новые подходы к разработке вакцин и использованию опосредованного паразитами подавления иммунитета для лечения воспалительных заболеваний 33 .
1-4). Локе и др. просмотреть последнюю информацию об иммунном ответе человека на паразитических гельминтов, узнать, как последние технологические разработки, такие как исследования на людях, секвенирование отдельных клеток и системы культивирования органоидов, помогли этим исследованиям. Недавние открытия позволяют использовать новые подходы к разработке вакцин и использованию опосредованного паразитами подавления иммунитета для лечения воспалительных заболеваний 33 .
Паразитические гельминты будут по-прежнему оставаться уникальным и физиологически значимым испытательным полигоном для новых концепций иммунологии слизистых оболочек. Срочно необходимо лучшее понимание иммунитета к гельминтам. Почти четверть населения мира подвержена риску заражения паразитическими гельминтами, вызывающими значительную заболеваемость и смертность, и все же у нас в настоящее время нет эффективных вакцин. Но, как показывает этот сборник обзоров и текущих исследовательских работ, понимание биологии гельминтозов, их регуляции и тканевого контекста также отвечает на ключевые вопросы иммунобиологии, иммунитета слизистых оболочек и иммунотерапии. Ответы на эти фундаментальные вопросы можно получить только с помощью экспериментов по заражению гельминтами на моделях человека и животных.
Ссылки
Ogilvie, B.M. Реагин-подобные антитела у животных, иммунных к гельминтозам. Природа 204 , 91–92 (1964).
Артикул КАС пабмед Google Scholar
Кей, А. Б. Ранняя история эозинофилов. клин. Эксп. Аллергия 45 , 575–582 (2015).
Артикул КАС пабмед Google Scholar
Бастен А., Бойер М. Х. и Бисон П. Б. Механизм эозинофилии. I. Факторы, влияющие на реакцию эозинофилов крыс на Trichinella Spiralis. Дж. Эксп. Мед. 131 , 1271–1287 (1970).
Артикул КАС пабмед ПабМед Центральный Google Scholar
 «>
«>Келли, Дж. Д. и Огилви, Б. М. Количество тучных клеток и эозинофилов в кишечнике во время изгнания червей у нерожавших и кормящих крыс, инфицированных Nippostrongylus brasiliensis. Междунар. Арка Аллергия Appl. Иммунол. 43 , 497–509 (1972).
Артикул КАС пабмед Google Scholar
Mosmann, T.R. & Coffman, R.L. Клетки Th2 и Th3: различные модели секреции лимфокинов приводят к различным функциональным свойствам. год. Преподобный Иммунол. 7 , 145–173 (1989).
Артикул КАС пабмед Google Scholar
Finkelman, F.D. et al. Опосредованная интерлейкином-4 и интерлейкином-13 защита хозяина от кишечных паразитов-нематод. Иммунол. Ред. 201 , 139–155 (2004).
Артикул КАС пабмед Google Scholar
 «>
«>Kuhn, R., Rajewsky, K. & Muller, W. Получение и анализ мышей с дефицитом интерлейкина-4. Наука 254 , 707–710 (1991).
Артикул КАС пабмед Google Scholar
Noben-Trauth, N. et al. Независимый от интерлейкина 4 (ИЛ-4) путь продукции ИЛ-4 CD4+ Т-клетками обнаружен у мышей с дефицитом рецептора ИЛ-4. Проц. Натл акад. науч. США 94 , 10838–10843 (1997).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Bancroft, A.J., McKenzie, A.N. & Grencis, R.K. Критическая роль IL-13 в устойчивости к кишечной нематодной инфекции. J. Иммунол. 160 , 3453–3461 (1998).
КАС пабмед Google Scholar
Cliffe, L. J. et al. Ускоренный оборот эпителиальных клеток кишечника: новый механизм изгнания паразитов.
 Наука 308 , 1463–1465 (2005).
Наука 308 , 1463–1465 (2005).Артикул КАС пабмед Google Scholar
Энтони, Р. М., Рутицкий, Л. И., Урбан, Дж. Ф. Младший, Стадекер, М. Дж. и Гаузе, В. К. Защитные иммунные механизмы при гельминтной инфекции. Нац. Преподобный Иммунол. 7 , 975–987 (2007).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Миллер, Х. Р., Хантли, Дж. Ф. и Уоллес, Г. Р. Иммунное отторжение и улавливание слизи во время быстрого изгнания Nippostrongylus brasiliensis из примированных крыс. Иммунология 44 , 419–429 (1981).
КАС пабмед ПабМед Центральный Google Scholar
Loke, P., MacDonald, A.S., Robb, A., Maizels, R.M. & Allen, J.E. Альтернативно активированные макрофаги, индуцированные нематодной инфекцией, ингибируют пролиферацию посредством межклеточного контакта.
 евро. Дж. Иммунол. 30 , 2669–2678 (2000).
евро. Дж. Иммунол. 30 , 2669–2678 (2000).Артикул КАС пабмед Google Scholar
Аллен, Дж. Э. и Винн, Т. А. Эволюция иммунитета Th3: быстрый восстановительный ответ на патогены, разрушающие ткани. ПЛОС Патог. 7 , e1002003 (2011 г.).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Jenkins, S.J. et al. Местная пролиферация макрофагов, а не рекрутирование из крови, является признаком воспаления Th3. Наука 332 , 1284–1288 (2011).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Глатман Зарецкий А. и др. Т-фолликулярные хелперные клетки дифференцируются из клеток Th3 в ответ на антигены гельминтов. Дж. Эксп. Мед. 206 , 991–999 (2009).

Артикул пабмед Google Scholar
King, I. L. & Mohrs, M. IL-4-продуцирующие CD4+ Т-клетки в реактивных лимфатических узлах при заражении гельминтами являются Т-фолликулярными хелперными клетками. Дж. Эксп. Мед. 206 , 1001–1007 (2009 г.).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Neill, D. R. et al. Нуоциты представляют собой новый врожденный эффекторный лейкоцит, который опосредует иммунитет типа 2. Природа 464 , 1367–1370 (2010).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Gerbe, F. et al. Клетки кишечного эпителиального пучка инициируют иммунитет слизистой оболочки 2 типа к паразитам-гельминтам. Природа 529 , 226–230 (2016).

Артикул КАС пабмед Google Scholar
Варьяни, Ф. и др. IL-25-зависимая цепь пучковых клеток, управляемая кишечными гельминтами, нуждается в факторе, ингибирующем миграцию макрофагов (MIF). Иммунол слизистых оболочек. (2022).
Котас, М. Э. и др. CISH ограничивает цепь tuft-ILC2, чтобы установить эпителиальный и иммунный тонус. Иммунол слизистых оболочек. 14 , 1295–1305 (2021).
Артикул КАС пабмед ПабМед Центральный Google Scholar
Инклан-Рико, Дж. М., Росси, Х. Л. и Герберт, Д. Р. «Каждая клетка является иммунной клеткой; вклад некроветворных клеток в противогельминтный иммунитет». Иммунол слизистых оболочек. (2022).
Spalinger, M. R. et al. Потеря протеинтирозинфосфатазы, не являющейся рецептором типа 2, снижает альтернативную активацию макрофагов, управляемую IL-4.
 Иммунол слизистых оболочек. 15 , 74–83 (2022).
Иммунол слизистых оболочек. 15 , 74–83 (2022).Артикул КАС пабмед Google Scholar
Паскаль, М. и др. Нейропептид VIP потенцирует кишечный врожденный иммунитет 2-го и 3-го типов в ответ на кормление. Иммунол слизистых оболочек. (2022).
Michla, M. & Wilhelm, C. Пища для размышлений – метаболизм ILC в контексте гельминтозов. Слизистая Иммунол. https://doi.org/10.1038/s41385-022-00559-у.
Мойат, М. и др. Микробная регуляция перистальтики кишечника обеспечивает устойчивость к гельминтозам. Иммунол слизистых оболочек. (2022).
Капсе, Б. и др. Зависимое от возраста повышение компетентности IFN-гамма подрывает эффективные ответы типа 2 на инфекцию нематодами. Иммунол слизистых оболочек. (2022).
Webster, H.
 C. et al. Тканевая передача сигналов IL-10 при гельминтозах ограничивает экспрессию IFNgamma и способствует кишечному Th3-ответу. Иммунол слизистых оболочек. (2022).
C. et al. Тканевая передача сигналов IL-10 при гельминтозах ограничивает экспрессию IFNgamma и способствует кишечному Th3-ответу. Иммунол слизистых оболочек. (2022).Classon, C.H. et al. Инфицирование кишечными гельминтами трансформирует CD4(+) Т-клеточный состав кожи. Иммунол слизистых оболочек. 15 , 257–267 (2022).
Артикул КАС пабмед Google Scholar
Вакка Ф. и Ле Гро Г. Тканеспецифический иммунитет при гельминтозах. Иммунол слизистых оболочек. (2022).
Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M. & Toda, M. Иммунологическая самопереносимость, поддерживаемая активированными Т-клетками, экспрессирующими альфа-цепи рецептора IL-2 (CD25). Нарушение единого механизма самопереносимости вызывает различные аутоиммунные заболевания. Дж. Иммунол. 155 , 1151–1164 (1995).

КАС пабмед Google Scholar
Майзелс, Р. М. и Язданбахш, М. Иммунная регуляция паразитами-гельминтами: клеточные и молекулярные механизмы. Нац. Преподобный Иммунол. 3 , 733–744 (2003).
Артикул КАС пабмед Google Scholar
Локе П., Ли С. К. и Ойесола О. О. Влияние гельминтов на иммунный ответ человека и микробиом. Иммунол слизистых оболочек. (2022).
Ссылки для скачивания
Информация об авторе
Авторы и организации
Школа инфекций и иммунитета Университета Глазго, Глазго, Великобритания
Georgia Perona-Wright
Отделение клеточной сигнализации и иммунологии, Школа наук о жизни, Университет Данди, Данди, Великобритания
Henry J. McSorley
Авторы
5
Вы также можете искать этого автора в PubMed Google Scholar
